
Glutamate is widely recognized as both a proteinogenic amino acid and the principal excitatory neurotransmitter in the central nervous system. In experimental biology, however, its significance extends well beyond either role. Glutamate connects amino acid interconversion, nitrogen transfer, the glutamine-glutamate-alpha-ketoglutarate axis, GABA synthesis, glutathione-dependent redox control, and tricarboxylic acid (TCA) cycle activity. This network position makes glutamate metabolism relevant to neuroscience, cancer biology, ferroptosis research, and systems-level metabolic analysis.
A glutamate concentration change is therefore not mechanistically specific by itself. Its interpretation depends on the biological matrix, cell type, subcellular compartment, sample handling, analytical method, and the behavior of related metabolites. This article outlines the major pathways and disease contexts of glutamate and explains how targeted LC-MS/MS, untargeted metabolomics, stable-isotope tracing, and multi-omics integration can address different research questions.
1. What Is Glutamate? From Amino Acid to Metabolic Hub
Glutamate has a straightforward chemical identity but a broad biological footprint. The terms glutamic acid, L-glutamate, and glutamate are sometimes used interchangeably in general writing, although they refer to different chemical descriptions. At physiological pH, the deprotonated form, glutamate, predominates.
1.1 Glutamic Acid, L-Glutamate, and Basic Biological Identity
Glutamic acid is an alpha-amino acid and one of the 20 standard amino acids incorporated into proteins. Its side-chain carboxyl group is negatively charged under most physiological conditions, which places it among the acidic amino acids. In humans, glutamate is generally considered non-essential because it can be produced through transamination reactions involving alpha-ketoglutarate and other amino acids.
This synthetic flexibility gives glutamate functions that extend beyond protein structure. It receives and donates amino groups, contributes to the synthesis of other amino acids and neurotransmitters, and links cytosolic reactions with mitochondrial metabolism. Consequently, glutamate is frequently included in amino acid panels, central carbon metabolism studies, and redox-focused metabolomics.
1.2 Major Biological Roles of Glutamate
The research value of glutamate comes from its participation in several biological layers. It is a protein building block, a central nitrogen carrier, a precursor of GABA and glutathione, and a source of alpha-ketoglutarate for the TCA cycle. In the nervous system, it activates ionotropic and metabotropic receptors to support synaptic communication. These functions are interconnected rather than independent, which is why pathway-level measurements are often more informative than a single-analyte result.
Table 1. Major Biological Functions of Glutamate
| Biological role | Primary context | Related molecules | Research relevance |
|---|---|---|---|
| Proteinogenic amino acid |
Protein and peptide synthesis |
Proteins, peptides |
Basic amino acid metabolism and biosynthesis |
|
Nitrogen carrier |
Transamination and ammonia handling | Aspartate, alanine, ammonia | Amino acid interconversion and nitrogen redistribution |
|
Carbon-metabolism intermediate |
Mitochondrial metabolism and TCA cycle | Glutamine, alpha-ketoglutarate | Anaplerosis, energy metabolism, and biosynthetic precursor supply |
|
Neurotransmitter and precursor |
Central nervous system | AMPA, NMDA, metabotropic receptors, GABA | Synaptic transmission, plasticity, and excitation-inhibition balance |
|
Redox-related precursor |
Glutathione metabolism | Cysteine, glycine, GSH, GSSG | Antioxidant defense and ferroptosis-related biology |
2. Central Pathways in Glutamate Metabolism
Glutamate metabolism is organized as a connected network rather than a sequence of isolated reactions. The central routes include glutamine conversion to glutamate, glutamate conversion to alpha-ketoglutarate, transamination, GABA production, glutathione synthesis, and transporter-dependent exchange between cells and compartments.
2.1 The Glutamine–Glutamate–Alpha-Ketoglutarate Axis
Glutamine is converted to glutamate by glutaminase, and glutamate can then be converted to alpha-ketoglutarate through glutamate dehydrogenase or aminotransferase reactions. Alpha-ketoglutarate enters the TCA cycle, linking amino acid metabolism with energy production, anaplerosis, and biosynthetic precursor generation.
Glutamate is therefore not simply an end product of glutamine catabolism. Its carbon skeleton and amino group can be redistributed through multiple reactions. Aspartate aminotransferase and alanine aminotransferase, for example, couple glutamate metabolism to aspartate and alanine production. The relative importance of these routes varies with tissue type, nutrient availability, oxygen status, and cellular demand.
2.2 Links to GABA, Glutathione, and Nitrogen Balance
Glutamate is the immediate precursor of gamma-aminobutyric acid (GABA), which is produced by glutamate decarboxylase. It is also incorporated into glutathione through a two-step pathway: glutamate-cysteine ligase first joins glutamate and cysteine, and glutathione synthetase then adds glycine. These reactions connect glutamate availability with inhibitory neurotransmission and antioxidant capacity.
Transamination provides a third major function. Glutamate can accept amino groups from other amino acids or donate its amino group to keto acids, thereby coordinating nitrogen redistribution across the amino acid pool. Changes in glutamate can therefore arise from altered carbon metabolism, neurotransmitter synthesis, glutathione demand, or nitrogen handling rather than from one pathway alone.
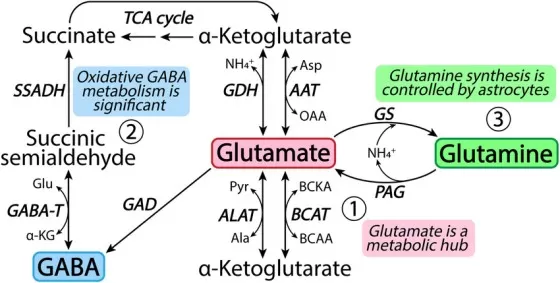
Figure 1. Glutamate as a metabolic hub connecting glutamine synthesis, alpha-ketoglutarate metabolism, the GABA shunt, and the TCA cycle. Reproduced from Andersen et al. (2021), Neuropharmacology, 196, 108719, under the Creative Commons Attribution 4.0 International License (CC BY 4.0).
2.3 Transporters and Cell-Type-Specific Interpretation
Transport processes strongly shape glutamate biology. Excitatory amino acid transporters remove extracellular glutamate in the nervous system and help maintain synaptic homeostasis. In redox and cancer biology, the cystine/glutamate antiporter SLC7A11/xCT exchanges intracellular glutamate for extracellular cystine, thereby supporting cysteine availability for glutathione synthesis (Koppula et al., 2021).
The meaning of a glutamate measurement also depends on the compartment sampled. Extracellular glutamate, intracellular glutamate, mitochondrial glutamate, and whole-tissue glutamate represent different biological pools. Plasma, cerebrospinal fluid, brain tissue, tumor tissue, cultured cells, and fecal material should not be interpreted using the same assumptions because cellular composition, transport, diet, microbial metabolism, and sample processing differ substantially across matrices.
3. Glutamate in the Nervous System
The nervous system is the best-known context for glutamate research. Its signaling role depends on receptor activation, rapid clearance from the extracellular space, and metabolic recycling between neurons and glial cells. Disruption at any of these levels can alter excitation-inhibition balance and cellular stress responses.
3.1 Glutamate Receptors and Excitation-Inhibition Balance
Glutamate acts through ionotropic receptors, including AMPA, NMDA, and kainate receptors, as well as metabotropic glutamate receptors. Ionotropic receptors are ligand-gated ion channels with distinct subunit compositions and physiological functions (Hansen et al., 2021). AMPA receptors support rapid excitatory transmission, whereas NMDA receptors contribute to calcium signaling, synaptic plasticity, and activity-dependent circuit remodeling.
Receptor-mediated signaling is closely linked to glutamate metabolism. The extracellular glutamate concentration must be tightly controlled, and neurotransmitter pools must be replenished without allowing prolonged receptor activation. GABA production from glutamate further connects glutamatergic metabolism with inhibitory signaling, making excitation-inhibition balance a network property rather than a single-metabolite phenomenon.
3.2 The Glutamate–Glutamine/GABA Cycle in Neuron–Glia Metabolism
After synaptic release, glutamate is taken up primarily by astrocytic transporters and converted to glutamine by glutamine synthetase. Glutamine can then be released, taken up by neurons, and converted back to glutamate by phosphate-activated glutaminase. In GABAergic neurons, glutamate can be decarboxylated to GABA. Together, these reactions support neurotransmitter recycling and metabolic communication between neurons and glial cells (Andersen, 2025).
Changes in astrocyte abundance, transporter activity, glutamine synthetase, neuronal glutaminase, or GABA synthesis can all influence measured glutamate. Recent work has also emphasized that dysfunction of the glutamate-glutamine cycle can accompany neurodegenerative processes and altered neuron-astrocyte communication (Sidoryk-Węgrzynowicz et al., 2024).
3.3 Excitotoxicity and Neurological Disease Interpretation
Excitotoxicity describes neuronal injury associated with excessive glutamate receptor activation and downstream calcium-dependent stress. It has been investigated in acute brain injury, ischemia, neuroinflammation, and neurodegenerative disease. Synaptic glutamate homeostasis depends on coordinated neuronal and astrocytic uptake and metabolism, and disruption of these processes can contribute to pathological signaling (Andersen et al., 2021).
An elevated glutamate concentration alone does not establish excitotoxicity. Stronger evidence requires concordant changes in receptor or transporter function, glutamine and GABA metabolism, oxidative stress, mitochondrial function, cellular injury, or physiological outcomes. In research reporting, glutamate is best described as an excitotoxicity-related readout unless direct functional evidence supports a causal claim.
4. Glutamate in Cancer Metabolism and Redox Biology
Outside the nervous system, glutamate is particularly important in cancer metabolism and cellular redox regulation. Its interpretation depends on how glutamine use, alpha-ketoglutarate production, transamination, cystine exchange, and glutathione synthesis are remodeled in the experimental system.
4.1 Glutamate in Cancer Metabolism
Cancer cells can remodel glutamine and glutamate metabolism to support proliferation, survival, and stress adaptation. Glutamine-derived glutamate can supply alpha-ketoglutarate for the TCA cycle and provide nitrogen for amino acid and nucleotide synthesis. Glutaminase activity, aminotransferases, glutamate dehydrogenase, nutrient transport, and the tumor microenvironment all influence the resulting metabolic phenotype (Jin et al., 2023).
The biological meaning of glutamate varies across tumors and model systems. A decrease in intracellular glutamate may reflect reduced glutaminolysis, increased export through SLC7A11, enhanced transamination, or altered mitochondrial entry. Conversely, accumulation may indicate increased glutamine catabolism, restricted downstream utilization, or changes in cell composition. Stable-isotope tracing is often required when the question concerns nutrient contribution or pathway flux rather than steady-state abundance.
4.2 SLC7A11, Glutathione, and Ferroptosis-Related Biology
Glutamate is linked to antioxidant defense through both glutathione synthesis and cystine transport. SLC7A11/xCT imports cystine in exchange for glutamate, providing a major source of cysteine for glutathione biosynthesis. High SLC7A11 activity can support redox defense and suppress ferroptosis, although it can also create context-dependent metabolic liabilities (Koppula et al., 2021).
Ferroptosis is a regulated cell-death process associated with iron-dependent lipid peroxidation and insufficient antioxidant protection. Glutamate-focused ferroptosis studies should therefore combine glutamate with cystine or cysteine-related metabolites, GSH and GSSG, lipid-peroxidation readouts, and relevant genes or proteins such as SLC7A11, GPX4, GCLC, and GCLM. This design separates a general amino acid change from evidence of redox or ferroptosis-related pathway remodeling.
Across cancer and redox studies, glutamate is most informative as part of a question-specific panel. The panel should reflect the hypothesized mechanism and the sample matrix rather than applying the same analyte list to every disease model.
4.3 Glutamate as a Disease-Related Metabolic Readout
Across disease models, glutamate becomes more informative when it is placed within a small pathway panel. Neuroscience studies often need glutamine, GABA, and transporter-related markers to separate neurotransmitter cycling from broader tissue injury. Cancer metabolism studies gain context from glutamine, alpha-ketoglutarate, aspartate, and TCA intermediates, which help distinguish anaplerosis, biosynthesis, and redox adaptation. Oxidative stress studies usually require glutathione, GSSG, cysteine, glycine, NADPH-related biology, and lipid peroxidation markers before a glutamate change can be linked to redox mechanisms.
Table 2. Glutamate-Related Readouts in Disease Research
| Research area | Key readouts | Interpretation |
|---|---|---|
| Neuroscience | Glutamate, glutamine, GABA | Neurotransmission, neurotransmitter recycling, excitotoxicity-related mechanisms |
| Cancer metabolism | Glutamine, glutamate, alpha-ketoglutarate, aspartate | Anaplerosis, metabolic reprogramming, biosynthesis, tumor stress adaptation |
| Redox biology | Glutamate, cysteine, glycine, GSH, GSSG | Oxidative stress and ferroptosis-related biology |
| Gut microbiome research | Luminal glutamate, amino acid metabolites, microbial pathway signals | Host–microbiome metabolic signaling and nutritional metabolism |
| Drug response studies | Glutamate pathway metabolites, transporter expression, pathway enzymes | Mechanism validation and treatment-induced metabolic remodeling |
This pathway-level approach is also more appropriate for biomarker discovery. A single metabolite may provide an initial signal, but a defensible biomarker claim still depends on reproducible measurement, independent validation, biological interpretability, and appropriate statistical modeling. For glutamate-related studies, the strongest conclusions usually come from consistent metabolite patterns rather than from glutamate alone.
5. LC-MS and Multi-Omics Strategies for Glutamate Research
Glutamate is analytically accessible but biologically context-sensitive. Platform selection should be based on whether the study requires validated concentration measurements, broad pathway discovery, metabolic-flux information, or integration with regulatory molecular layers.
5.1 Targeted LC-MS/MS for Quantitative Analysis
Targeted LC-MS/MS is appropriate when predefined analytes, reproducibility, and concentration-level interpretation are required. A robust workflow may include authentic standards, isotope-labeled internal standards where available, calibration curves, pooled quality-control samples, and matrix-specific validation. These measures are important because ion suppression, recovery differences, fragmentation, isomers, and peak misidentification can affect metabolomics results (Alseekh et al., 2021).
Glutamate and glutamine are highly polar, so chromatographic retention, derivatization strategy, matrix effects, and separation from related compounds should be considered during method development. Absolute quantification should be claimed only when the calibration and validation strategy supports it. Otherwise, the result should be described as relative or semi-quantitative.
5.2 Untargeted Metabolomics for Broader Pathway Discovery
Untargeted metabolomics can identify wider metabolic perturbations surrounding glutamate, including changes in amino acids, organic acids, nucleotides, redox-associated metabolites, and lipid metabolism. It is useful when the pathway of interest is not fully defined or when unexpected metabolic responses are important.
Untargeted results require careful annotation. Exact mass and retention behavior alone do not provide the same confidence as comparison with an authentic standard and matched MS/MS data. Reporting should distinguish confirmed metabolites from putative annotations, and biological conclusions should reflect the annotation level and analytical coverage (Alseekh et al., 2021).
5.3 Multi-Omics Integration for Mechanism-Oriented Interpretation
Multi-omics integration can connect metabolite changes with genes, proteins, transporters, and signaling pathways. A glutamate-centered design may combine glutamine, glutamate, GABA, alpha-ketoglutarate, aspartate, GSH, and GSSG with glutaminase (GLS), glutamate dehydrogenases (GLUD1/2), glutamine synthetase (GLUL), glutamate decarboxylases (GAD1/2), glutamate-cysteine ligase subunits (GCLC/GCLM), the glutamine transporter SLC1A5/ASCT2, excitatory amino acid transporters such as SLC1A2/EAAT2 and SLC1A3/EAAT1, and SLC7A11/xCT.
Transcriptomics can identify changes in pathway gene expression, whereas proteomics more directly measures enzyme and transporter abundance. Phosphoproteomics may add information when pathway activity is regulated post-translationally. Cross-omics agreement can strengthen a mechanistic hypothesis, but integrated association alone does not constitute mechanism validation; functional perturbation or orthogonal experiments are still required for causal conclusions.
5.4 Common Analytical and Biological Pitfalls
Pre-analytical handling can substantially affect polar metabolites. Delayed quenching, temperature variation, repeated freeze-thaw cycles, extraction conditions, and storage can alter measured glutamate or glutamine. Internal standards intended to correct extraction recovery should generally be added before or at the start of extraction, while pooled QC samples should be distributed across the analytical sequence to monitor stability and drift.
Biological interpretation can also be confounded by cell composition, diet, medication, microbiome activity, tissue injury, and matrix effects. Statistical significance does not by itself establish pathway direction or causality. The most reliable conclusions combine a prespecified biological question, appropriate controls, matrix-matched analytical validation, related metabolites, and orthogonal biological evidence.
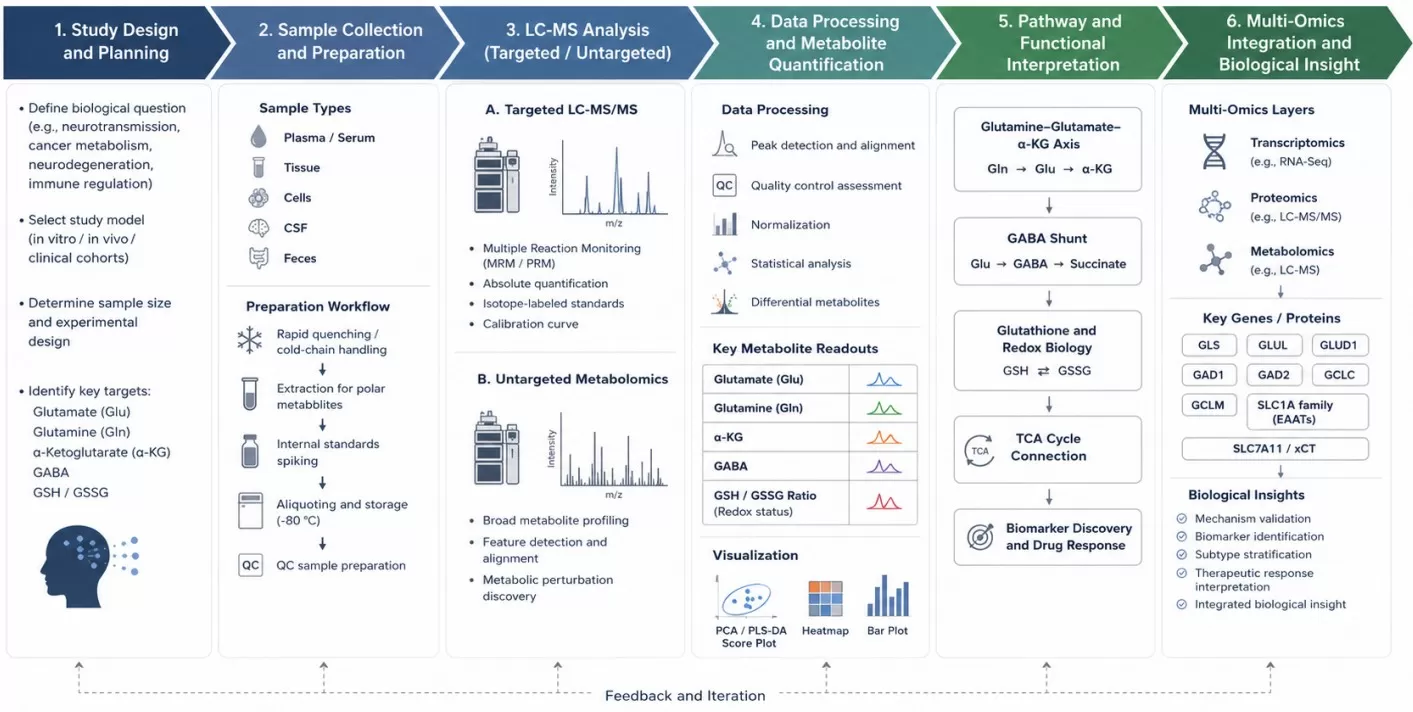
Figure 2. Workflow for glutamate-focused LC-MS metabolomics and multi-omics integration, from study design and sample handling to targeted quantification, pathway analysis, and gene/protein integration.
6. Building a Glutamate-Focused Omics Study with MetwareBio
A useful glutamate study begins with the biological question rather than the platform. Neuroscience projects may prioritize glutamate, glutamine, GABA, and transporter-related interpretation. Cancer metabolism projects may focus on glutamine use, alpha-ketoglutarate formation, aspartate and TCA-cycle metabolism, or isotope tracing. Redox and ferroptosis studies may require glutamate to be measured together with cysteine-related metabolites, GSH/GSSG, lipid-peroxidation markers, and pathway proteins.
MetwareBio supports question-driven study design through amino acid targeted metabolomics, untargeted metabolomics, and multi-omics integration. Targeted assays can provide concentration-level information for predefined amino acids and related metabolites, whereas untargeted profiling broadens pathway coverage. Transcriptomic and proteomic layers can be added when the study requires regulatory context or mechanism-oriented interpretation.
The final workflow should be matched to the sample type, expected effect size, study model, sample number, analytical validation requirements, and the strength of the biological claim. This planning step reduces overinterpretation and ensures that each dataset contributes directly to the research objective.
Planning a glutamate metabolism study? Contact MetwareBio to discuss a suitable targeted, untargeted, or multi-omics workflow.
Contact UsRead More: Glutamate Metabolism and Multi-Omics Research
Explore these related articles to deepen your understanding of glutamate metabolomics, from amino acid quantification and neurotransmitter analysis to TCA cycle connections and multi-omics integration strategies.
Glutamate sits at the center of amino acid metabolism. Learn how targeted amino acid metabolomics quantifies glutamate alongside glutamine, GABA, aspartate, and other pathway-relevant amino acids to support robust pathway-level interpretation in disease research.
As the major excitatory neurotransmitter, glutamate is central to neuroscience metabolomics. This article explains how neurotransmitter-targeted metabolomics measures glutamate, GABA, and related signaling molecules in brain tissue, CSF, and plasma for neuroscience studies.
This article compares targeted, untargeted, and widely-targeted metabolomics approaches, helping you choose the right analytical strategy for glutamate research based on your research question, sample type, and pathway coverage requirements.
Glutamate connects to the TCA cycle through alpha-ketoglutarate. This article explains TCA cycle biology and its metabolic connections, providing the pathway context needed to interpret glutamate changes in cancer metabolism and energy research.
This article explains how combining proteomics with metabolomics connects glutamate abundance with enzyme expression, transporter levels, and pathway activity, supporting the multi-omics integration approach discussed for mechanistic interpretation.
For researchers planning a glutamate-focused multi-omics study, this overview covers the advantages and application areas of multi-omic analysis, from study design to integrated data interpretation across disease research contexts.
References
- Alseekh, S., Aharoni, A., Brotman, Y., Contrepois, K., D’Auria, J., Ewald, J., Ewald, J. C., Fraser, P. D., Giavalisco, P., Hall, R. D., Heinemann, M., Link, H., Luo, J., Neumann, S., Nielsen, J., Perez de Souza, L., Saito, K., Sauer, U., Schroeder, F. C., … Fernie, A. R. (2021). Mass spectrometry-based metabolomics: A guide for annotation, quantification and best reporting practices. Nature Methods, 18, 747–756. DOI: 10.1038/s41592-021-01197-1
- Andersen, J. V. (2025). The glutamate/GABA-glutamine cycle: Insights, updates, and advances. Journal of Neurochemistry, 169(3), e70029. DOI: 10.1111/jnc.70029
- Andersen, J. V., Markussen, K. H., Jakobsen, E., Schousboe, A., Waagepetersen, H. S., Rosenberg, P. A., & Aldana, B. I. (2021). Glutamate metabolism and recycling at the excitatory synapse in health and neurodegeneration. Neuropharmacology, 196, 108719. DOI: 10.1016/j.neuropharm.2021.108719
- Hansen, K. B., Wollmuth, L. P., Bowie, D., Furukawa, H., Menniti, F. S., Sobolevsky, A. I., Swanson, G. T., Swanger, S. A., Greger, I. H., Nakagawa, T., McBain, C. J., Jayaraman, V., Low, C. M., Dell’Acqua, M. L., Diamond, J. S., Camp, C. R., Perszyk, R. E., Yuan, H., & Traynelis, S. F. (2021). Structure, function, and pharmacology of glutamate receptor ion channels. Pharmacological Reviews, 73(4), 1469–1658. DOI: 10.1124/pharmrev.120.000131
- Jin, J., Byun, J.-K., Choi, Y.-K., & Park, K.-G. (2023). Targeting glutamine metabolism as a therapeutic strategy for cancer. Experimental & Molecular Medicine, 55, 706–715. DOI: 10.1038/s12276-023-00971-9
- Koppula, P., Zhuang, L., & Gan, B. (2021). Cystine transporter SLC7A11/xCT in cancer: Ferroptosis, nutrient dependency, and cancer therapy. Protein & Cell, 12(8), 599–620. DOI: 10.1007/s13238-020-00789-5
- Sidoryk-Węgrzynowicz, M., Adamiak, K., & Strużyńska, L. (2024). Astrocyte-neuron interaction via the glutamate-glutamine cycle and its dysfunction in tau-dependent neurodegeneration. International Journal of Molecular Sciences, 25(5), 3050. DOI: 10.3390/ijms25053050